Estuaries: Where the Land Meets the Sea
Bonnethead Shark
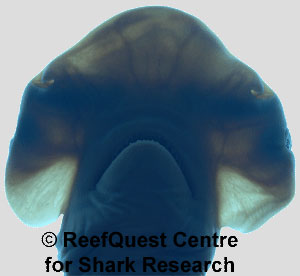
With its modestly-developed, spade-shaped head, the Bonnethead (Sphyrna tiburo) is less overtly weird than other hammerheads and is actually rather . . . well, cute. But no matter how cute they may seem to us, Bonnetheads are exquisitely-adapted and in their own, small-scale way socially complex and formidable predators in warm shallow seas and estuaries.
Just the Facts:
Size:
Reproduction:
Diet:
Habitat: Intertidal, Estuaries, Sandy Plains, Fresh Waters Depth: Intertidal to 260 ft (80 m) Distribution: Tropical Eastern Pacific, Western North Atlantic, Caribbean, Amazonian |

A six-month study of a colony of adolescent Bonnetheads in a semi-natural enclosure at the Miami Seaquarium revealed that this species has surprisingly complex behavior, body language and social organization. No fewer than 18 Bonnethead body postures and movement patterns were identified, almost half of which appeared to regulate social activities. These included head shakes, back hunching, puffing out the gill pouches, jaw snapping, hitting another Bonnethead between the dorsal fins with the snout tip, circling head-to-tail, in lines of up to five Bonnethead following one another, and (in males) clasper flexing and thrusting. A subtle, size-related dominance hierarchy was also noted among the Bonnetheads, with submissive individuals giving way to dominants as little as 5 to 7.5% longer than themselves. This suggests that Bonnetheads have a keen awareness of their own size relative to that of others sharing their environment.
Bonnetheads are not only sensitive and responsive to one another, but also to dissolved chemicals in their environment. Due to the polar and ion-rich nature of seawater, dissolved chemicals disperse very quickly. Yet a series of experiments carried out on captive Bonnetheads revealed that they can actually detect which nare (nostril), the left or the right, receives the stronger whiff of a dissolved chemical. This suggests that simply having a modest hammer in which the nares are not separated by nearly the distance as those of larger, more grandly endowed hammerheads does not significantly hinder Bonnetheads in efficiently tracking water-borne odors.
If as several lines of evidence suggest enhanced sensory acuity is a major selective advantage of hammerheaded-ness in sharks, it seems that electroreception may be more important than olfaction in prey detection by hammerheads. The electrosensitive organs of sharks are termed ampullae of Lorenzini (after the Italian anatomist who first described them in 1678). As the name suggests, the ampullae are tiny, flask-like cells. With sensory hair cells at the base and filled with an electrically conductive jelly, each ampulla opens to the external environment via a small pore. These pores are concentrated over the undersurface of a sharks head, resembling a bad case of five oclock shadow. Compared with other hammerheads, the Bonnetheads ampullae are particularly richly distributed in symmetrical patterns over the entire undersurface of their hammers, including along the front margin, in front of the mouth, as well as under the nares and eyes. This suggests that electroreception may play a particularly important role in Bonnethead prey detection.
 A recent study explored the diet and feeding habits of Bonnethead Sharks
in estuarine waters of southwest Florida. Bonnethead stomach contents were
examined from two estuaries, Tampa Bay and Charlotte Harbor, and were found
to be very similar. In both locations, the diet of Bonnethead Sharks
consisted mostly of the commercially-important Blue Crab (Callinectes
sapidus) and of all things seagrasses.
A recent study explored the diet and feeding habits of Bonnethead Sharks
in estuarine waters of southwest Florida. Bonnethead stomach contents were
examined from two estuaries, Tampa Bay and Charlotte Harbor, and were found
to be very similar. In both locations, the diet of Bonnethead Sharks
consisted mostly of the commercially-important Blue Crab (Callinectes
sapidus) and of all things seagrasses.
That seagrasses are such a dominant component of Bonnethead diet is intriguing, because such grasses often feature in the stomach contents of many species of tropical, shallow-water sharks. It seems likely that the grass is consumed incidentally as the sharks feed on invertebrates and fishes hiding among the blades. Since vertebrates lack the enzymes necessary to digest cellulose, I suspect that ingested seagrass may act as roughage to help move food and indigestible bits such as crab external skeletons, which are composed of a hard, cellulose-like material called chitin through the intestinal valve. But this is pure speculation on my part. A colleague of mine thinks these sharks are actually digesting the grasses, as the cells appear to be in the process of breaking-down and stomach contents are often stained green. But I think that the acids secreted by the shark stomach are simply causing the plant cells to burst, allowing chlorophyll to leech out. Time and further investigation may shed more light on this mystery.
The dietary study also found that Bonnethead Shark feeding activity peaked in late afternoon and there is a direct relationship between shark length and Blue Crab carapace (upper shell) width, with longer sharks taking wider crabs. During the fall, Bonnethead Shark diet expanded to include two other crab species and at least one species of squid. This suggests that, like other sharks, Bonnetheads shift their diet to take advantage of seasonal availability of prey. Perhaps most intriguingly, although one would predict older sharks would have a slower metabolic rate and thus require less food per unit mass than younger ones, the study found that large Bonnetheads had proportionately more food in their stomachs than small ones. No one knows what to make of this result.

Another Bonnethead mystery concerns distinct geographic differences in birth size and growth rates. A recent series of studies has demonstrated that Bonnethead Sharks in Tampa Bay are born at a larger size and grow to maturity faster than those in the Florida Keys. Tampa Bay Bonnetheads are born at a length of about 14 inches (35 centimetres) and reach sexual maturity at an age of 24 to 29 months, while those from the Florida Keys are born at 11 inches (27 centimetres) and reach sexual maturity at 29 to 36 months.
Although the researchers declined to comment on why this pattern might be so clearly developed between these two Bonnethead populations, I suspect that newborn Bonnetheads might more vulnerable to predators in Tampa Bay than they are in the Florida Keys. If this is so, a likely culprit may be the Bull Shark (Carcharhinus leucas), a large species able to tolerate wide fluctuations in salinity and having a well-known predilection for dining on the pups of other shark species. Sport angling and commercial fishing records indicate that the Bull Shark is significantly more abundant in the northern Gulf of Mexico than in the Florida Keys. Further, the mangrove prop-roots and complex coral topography of the Florida Keys may provide some protection from would-be predators. Since swimming efficiency increases and vulnerability to predators decreases with size, larger pup size and faster growth rate would be highly beneficial to Tampa Bay Bonnetheads.
Whatever the reasons for these regional differences in Bonnethead birth size and growth rates, it is clear that estuarine waters are important nursery areas for this species. Shallow bays and mangrove swamps provide Bonnethead Sharks with shelter and food during the most vulnerable stage of their lives, increasing their likelihood of surviving to adulthood and beginning their species life cycle anew.
Bonnethead Shark Bibliography
More about: Function of the
Hammer | Origin and Evolution of the
Hammer
