No Guts, No Glory
Capturing prey no matter how spectacularly is not, of itself, enough. Neither is merely swallowing it. An animal such as a shark or a human with a two-opening gut (with a mouth at one end and a cloaca or anus at the other) can be thought of as a tube. By this analogy, a fish or seal in a White Shark's stomach is no more inside the shark than a finger inserted through the hole of a doughnut is inside the doughnut (those of you more health-conscious may substitute a bagel for the doughnut in this example the analogy holds). Somehow, food surrounded by the shark's stomach must be passed into the shark itself. Therefore, how the White Shark processes and absorbs a food item is thus vitally important to its ability to make use of the caloric energy it contains.
|
Gastrointestinal Tract of a
White Shark |
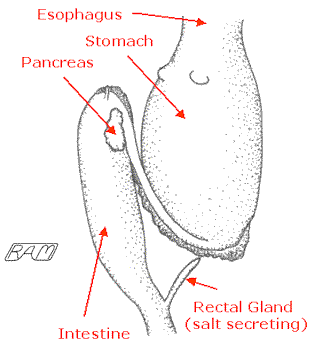
Like that of other sharks, the esophagus of the Great White is relatively short and lined with finger-like extensions that help prevent food escaping out the mouth. In the floor of the mouth is a small, relatively immobile 'tongue' (actually a forward-projecting extension of the cartilaginous supports at the midline of the gill 'basket' to which the hyoid arches attach). Control of food moving into the White Shark's stomach is accomplished by way of a contractile ring of muscle called a sphincter. The stomach itself is a or J-shaped organ, constituting about 20% of the shark's body length, and located along the midline of the body just posterior to the gills. The stomach walls are quite muscular, the inner lining having longitudinal folds called "rugae" which permit accordion-like expansion of the stomach to accommodate a particularly large meal. The maximum capacity of a White Shark's stomach is about 10% of its total weight, an ability which must come in handy for an opportunistic predator. In addition, the stomach lining is peppered with various secretory cells. Some of these cells produce hydrochloric acid (which helps soften and break-down bone), others copious amounts of mucus (to protect the stomach lining from digesting itself). In addition, the pancreas (which produces protein-splitting enzymes) has a duct that empties into the stomach, thereby beginning the process of protein break-down.
The stomach wall itself is largely composed of transverse rings of muscle, which through a coordinated series of rhythmic squeezings called "peristalsis" churn and mix the food and digestive secretions into a gloopy paste. The control of food out of the stomach and into the intestine is governed by another sphincter. Despite all its destructive machinery, the shark stomach can be used to store food for prolonged periods (on the order of months), somehow over-riding the stomach-distention sensor that would otherwise initiate the stomach's food break-down processes. As in humans, indigestible items can be voided out the mouth. But, unlike ourselves, at least some sharks can evert the stomach at will: turning it inside-out through the mouth, rinsing the stomach lining in sea water, then retracting and returning it to the normal inside-in condition. Although no one has yet reported this behavior in the Great White, sharks can apparently do this all about as easily as you or I might return an inside-out sock to its correct topology.
Another shark anatomical peculiarity is the liver. The liver of a shark is composed of two large lobes, left and right, and a much smaller median lobe. Unlike the dark chocolate-brown liver of the Spiny Dogfish (Squalus acanthias) a creature which many of us dissected as part of our formal education in high school and/or university the liver of the White Shark is typically a pale orangy-peach in color. Compared with other animals, the liver of sharks is very large typically accounting for 15 to 25% of the total body weight, and up to 35% of the total weight in some mesopelagic (mid-water) squaloids. Compare this with an average liver weight of about 1.5 to 2.2% of total body weight in humans. So, what do sharks in general and Great Whites in particular do with all that liver?
The vertebrate liver is a remarkable chemical factory, performing literally hundreds of roles in the body. Among many other functions, the liver banks vitamins for release in times of low supply, manufactures a starch-like compound that is used as a fuel supply by white muscle and can be used by other tissues in an emergency, stabilizes the body's blood-sugar level, detoxifies poisons, builds enzymes, processes digested fats, manufactures bile and cholesterol, and constitutes a major source of metabolic heat. In sharks, the liver is perfused with low-density oils and hydrocarbons. One of the most important of these hydrocarbons is squalene (C30H50), which is much less dense than seawater. Because sharks lack a swim bladder (an organ found in many teleosts that controls the overall density of the fish by the controlled secretion and absorption of the gases within it), sharks are heavier than water. The collective effect of the low-density compounds in the shark liver is to provide lift by reducing its overall density. As a result, a shark is only very slightly more dense than sea water, making a 'typical' shark only slightly heavier than the medium through which it swims. For example, an intriguing experiment by US Navy researcher H. David Baldridge revealed that a 1015-pound (461-kilogram) Tiger Shark (Galeocerdo cuvier) had an apparent weight in seawater of only 7.3 pounds (3.3 kilograms) a reduction of over 99%! Thanks largely to its oily liver, a shark must invest very little energy to prevent sinking and the vast majority of its swimming effort can go toward propulsion.
The shark liver thus acts as a kind of internal float, freeing these animals from depending solely on their foils to provide lift. It is widely stated that the liver also serves sharks as an energy store, as it does in most vertebrates a position Baldridge very much doubts, given that the last thing a starving shark should do is mess with its ability to swim. However, in a 1987 paper, South African zoologist G.J. Rossouw reported that the liver weight and oil content of the Lesser Guitarfish (Rhinobatos annulatus) increase dramatically during periods of peak mating activity. These changes could represent an energy store to fuel the muscles for this species' annual migration, or it could simply be a way to temporarily increase the Guitarfish's buoyancy while pupping or mating in brackish water (which is less dense, and thus less buoyant, than full-strength seawater). The White Shark has a relatively small liver by shark standards, typically constituting 8 to 12% of the total body weight. In general, there is an inverse relationship in sharks between activity level and liver size. Thus the White Shark's relatively small liver is consistent with its active lifestyle. This may mean that the White Shark is more dependent upon dynamic lift than, for example, the slow-swimming Basking Shark or a mesopelagic squaloid. Further, if there is any truth to the idea that sharks draw on energy stored in the liver during lean times (a notion for which there is some circumstantial evidence, but to date has only been conclusively demonstrated under conditions of extreme starvation), then the Great White may be more tightly bound to areas offering rich feeding than are other sharks.
In terms of energy per unit of mass, fats are rich food (which is why extra calories stored in human 'love handles' are composed of fat rather than protein: less weight to carry around). The relatively simple fats and oils produced by fishes are fairly easy to break-down, but the more complex fats that compose marine mammal blubber are much more difficult to digest. The liver produces a thick yellowish-green liquid called bile, which contains enzymes that emulsify fats. By reducing fat globules to smaller droplets, bile increases the amount of surface area of each droplet relative to its volume, thereby making it easier to digest. Bile from the liver is stored in the gall bladder, a greenish sac located on the underside of the median lobe. From there, it is transported (via a duct) to the anterior part of the intestine. The intestine is the complex tube at the posterior part of the vertebrate digestive tract where the actual absorption of nutrients occurs. Only after broken-down nutrients are passed through the intestinal wall are they inside the body and available to fuel its many processes.
|
|
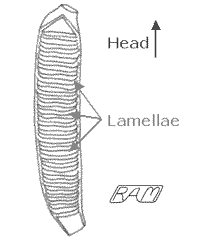
In an average-sized adult human, the combined length of the small and large intestine is about 26 feet (8 metres). Yet the intestine of a comparably-sized shark is only about a foot (30 centimetres) in length. Despite the intestine's shortness, it manages to absorb enough nutrients to maintain the shark due to a system of internal partitions. Known as intestinal valves, they are a triumph of compact packaging greatly increasing the internal surface area of the intestine without significantly increasing its length. In addition, these partitions greatly increase the efficiency of nutrient absorption even further by greatly increasing the time required for food to pass through the intestine. For example, shark biologist Brad Wetherbee and his co-workers used X-ray technology to measure the of the rate of food passage through the stomach and intestinal valve of juvenile Lemon Sharks fed on a uniform diet. They found that the sharks began initial voiding of undigested matter some 16 to 17 hours after feeding and that to completely empty the digestive tract required an astonishing 68 to 82 hours. In contrast, humans may process food and excrete solid waste in as little as four hours after eating (considerably less for certain spicy foods!)
There are three basic types of intestinal valve in sharks, termed spiral, scroll, and ring. The spiral valve is the 'classic' shark intestinal partition, resembling an auger in shape, and is found in cow sharks (family Hexanchidae), spiny dogfishes (Squalidae), and catsharks (Scyliorhinidae). The scroll valve resembles a loose roll of paper in shape, and is found in whaler sharks (Carcharhinidae, such as the Lemon Shark used in Wetherbee's gut-clearance experiment). The ring valve, resembling a series of tightly-packed lamellae (plates), is found in all extant lamnoids including the White Shark. The ring valve offers the most absorptive surface area per unit length, and is thus highly efficient. This efficiency is in-keeping with the White Shark's high-energy lifestyle. Further, the intestine in this species is typically about 10% as long as its body, roughly 25% as wide as it is long, and has from 47 to 54 lamellae ('rings'). For a 6-foot (2-metre) -long White Shark, that works out to a total absorptive surface area of about 570 to 660 square inches (3,690 to 4,240 square centimetres), or about the same as a Monopoly board. By comparison, the total surface area of the small and large intestines of a 6-foot-tall man is about 495 square inches (3,175 square centimetres) actually some 25% less than a comparably-sized White Shark.
But, just as a car produces exhaust, the White Shark does not use its fuel with 100% efficiency. After its ring valve has removed as much nutrient as possible from the stately parade of organic matter passing through the intestine, what solid matter that remains is excreted as waste. Because sharks do not have separate openings for products of the digestive and reproductive systems, the posterior opening of the gut is referred to as a cloaca instead of as an anus, as in humans. Rather than producing a more-or-less solid bolus of waste matter, as do human anuses (most of the time, anyway), shark cloacae void feces that is generally quite liquidy, typically a dirty mustard brown in color. Sometimes, a streaming cloud of shark feces is punctuated with an unexpectedly pretty surprise: a burst of silvery fish scales, glittering like diamonds in the flickering underwater light. With a graceful swish of its tail, the White Shark dissipates the momentary discoloration, returning its unused energy to the sea.
